−研究成果報告 2nd report−
2号連続で臼尻OBの最新研究論文の要約を掲載します。
第2回は、阿部拓三博士による、
Ichthyological Research, 54号 (2007)に掲載された論文です。
Histological structure of the male reproductive organs and spermatogenesis
in a copulating sculpin, Radulinopsis taranetzi (Scorpaeniformes: Cottidae)
(邦題:キマダラヤセカジカRadulinopsis taranetzi(カジカ科)雄における生殖器官系の構造と精子形成過程)
Takuzo Abe(1)* and Hiroyuki Munehara(2)
(1) Shizugawa Nature Center, 40 Sakamoto, Minamisanriku-cho Togura, Motoyoshi-gun,
Miyagi 986-0781, Japan
(2) Field Science Center for Northern Biosphere, Usujiri Fisheries Laboratory,
Hokkaido University, 152 Usujiri-cho, Hakodate, Hokkaido 041-1613, Japan
出典:Ichthyological Research 54: 137-144. (2007)
海産カジカ類には、交尾種(交尾型カジカ)と非交尾種がある。交尾型カジカでは、交尾によって精子は卵巣内へ輸送されるが、卵の胚発生が開始するのは、産卵によって海水と卵が接した瞬間である。この一風変わった受精様式は「体内配偶子会合」と呼ばれ、全ての海産交尾型カジカ共通の受精様式である。しかし、交尾型カジカの中でも、生殖器官系の構造・機能は種ごとに大きく異なり、それを反映して繁殖生態も多様である。
本研究では、2001年に新種記載された交尾型カジカ、キマダラヤセカジカの雄性生殖器官系の構造を組織学的に観察し、その機能形態について考察した。
供試魚は、函館市臼尻町沿岸で採集したキマダラヤセカジカの雄98個体を用いた。組織観察に基づく生殖器官系の発達過程と生殖腺重量指数 [GSI:
(精巣重量/体重)×100] の推移から以下の特徴が明らかとなった。
1. 生殖器官系は一対の精巣、貯精嚢、および生殖突起によって構成されていた(図1)。
2. 精子形成は、精巣後端部から前方に向かって順次進行し、精巣の成熟に部位による顕著な勾配が観察された(図2)。
3. 精子は、PAS反応に陽性を示すエオシン好染性物質とともに貯精嚢内に貯えられ、精漿内ではいかなる運動性も示さなかった(図3a-c)。
4. 精巣後端部では、生殖細胞包嚢の発達に伴って精小嚢内の上皮細胞が著しく肥厚し、精小嚢内腔は精子およびエオシン好染性物質によって満たされていた(図3d-f)。
5. 精子およびエオシン好染性物質が貯精嚢内に観察される期間は2-7月であった(表1)。
6. GSI値は、8-10月は低値(平均で0.0-0.3%)で推移するが、10月(0.3±0.10%)から12月上旬(9.6±2.29%)にかけて急激な増加を示した。しかし、1月には5.6±0.82%まで減少し、2月には3.8±0.63%となった。その後7月上旬(1.0±0.38%)まで緩やかな減少傾向を示し、8月には低値(0.3±0.04%)となった(図4)。
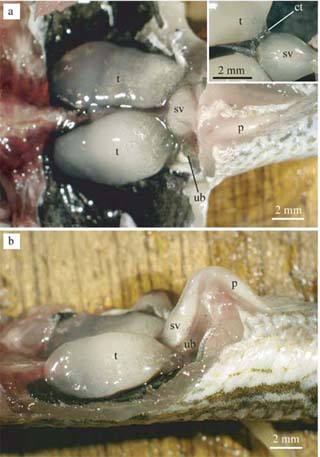
図1. キマダラヤセカジカの雄性生殖器官系.
a 腹側および精巣と貯精嚢をつなぐ
輸精管系の薄組織(挿入図).
b 横側. p: 生殖突起, t: 精巣, sd: 輸精管, ub: 膀胱 |
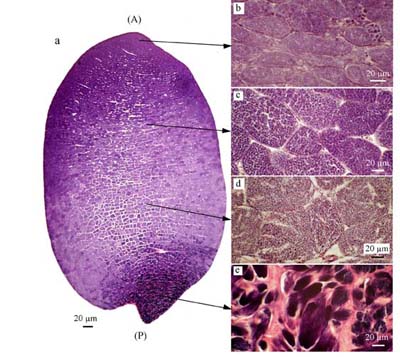
図2. キマダラヤセカジカの精巣(11月)の矢状面および各部位の拡大図.
(a)前端部(A)から後端部(P)にかけて
精子形成活性の顕著な勾配が見られる.
(b)精原細胞包嚢, (c)精母細胞包嚢,
(d)精細胞包嚢,(e)精子包嚢. |
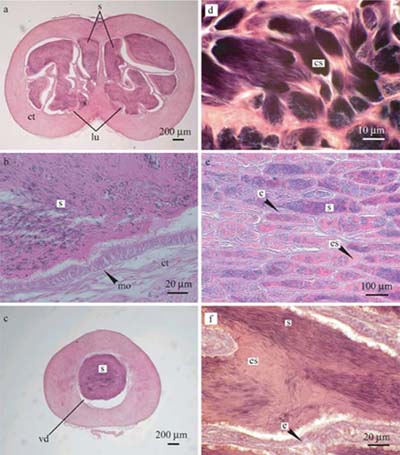
図3. キマダラヤセカジカの貯精嚢と精巣の組織図.
a 貯精嚢前中部の横断面図. b 貯精嚢内の上皮細胞. c 貯精嚢後部の横断面図. d精子包嚢.
e-f 精子およびエオシン好染生物質が充満した精小嚢内腔. cs: 精子包嚢, st: 結合組織,
e: 精小嚢内腔の上皮細胞, es: エオシン好染性物質, lu: 内腔, mo: 円柱上皮細胞, s: 精子, vd: 精管 |
表1. キマダラヤセカジカ雄の貯精嚢内に精子が貯留されていた頻度の季節変化.
各月の数字は、採集された全個体中に占める貯精嚢内での精子貯留が観察された個体数を示す.
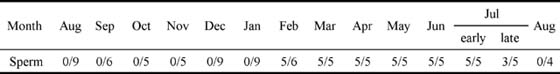 |
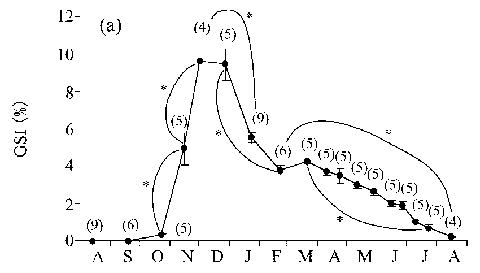
図4. キマダラヤセカジカ雄のGSIの季節変化.
括弧内はサンプル数. エラーバーは標準誤差.
*有意差(p<0.05; 各月の値についてone-way ANOVAでグループ間の差を検出し,
平均値の差をScheffe’s testを用いて検定). |
組織学的観察から、本種の生殖細胞は精巣後端部から順次成熟して放出され、エオシン好染性物質とともに貯精嚢内に供給されることが示された。また、精漿内における精子が運動活性を示さなかったことから、エオシン好染生物質は精子の運動を抑制する機能を持つ可能性が考えられる。GSI値における12月にみられた急激な減少は、1月に採集した個体の貯精嚢内に精子貯留が確認できなかったことから放精による精子消費によるものではなく、精子変態に伴う生殖細胞の染色質の凝集、および細胞質を残余小体として吸収することによると考えられる。以上から、本種の交尾期間は、精子およびエオシン好染性物質が貯精嚢内に観察される2-7月の6ヶ月に及ぶと考えられた。
本種の産卵期は5-7月に見られ、産卵期と交尾期のずれおよび交尾期間は他の交尾を行う魚類と比較しても著しく長期化している。また、カサゴ目魚類の他の交尾種では、精子は精漿内で活発な運動活性を示すことが知られている。本研究で観察された生殖器官系の発達様式および精子の運動抑制は、本種の長期化した交尾期間と強く関連した機能形態と考えられる。
表紙に戻る